
단백질생성성 아미노산

단백질생성성 아미노산(영어: proteinogenic amino acid)은 번역 과정에서 단백질의 생합성에 사용되는 아미노산이다. "단백질생성성(proteinogenic)"이라는 단어는 "단백질 생성(protein creating)"을 의미한다. 알려진 모든 생물체에서 22가지의 유전적으로 암호화된(단백질생성성) 아미노산이 있으며, 표준 유전 부호로 20가지, 특수 번역 메커니즘에 의해 통합될 수 있는 2가지(셀레노시스테인 및 피롤리신)이 있다.[1] 표준 아미노산(영어: common amino acid)이라고도 한다.
대조적으로 단백질비생성성 아미노산은 단백질에 통합되지 않거나(GABA, L-도파 또는 트라이아이오도티로닌과 같은) 유전적으로 암호화된 아미노산 대신 잘못 통합되거나 세포에 의해 직접적으로 분리되거나 생성되지 않는 아미노산(예: 하이드록시프롤린)이다. 후자는 보통 단백질의 번역 후 변형으로 인해 발생한다. 일부 단백질비생성성 아미노산은 비리보솜 펩타이드 합성효소에 의해 합성되는 비리보솜 펩타이드에 통합된다.
진핵생물과 원핵생물 모두 SECIS 요소로 알려진 뉴클레오타이드 서열을 통해 셀레노시스테인을 단백질에 통합시킬 수 있으며, 이는 세포가 근처의 UGA 코돈을 셀레노시스테인(UGA는 일반적으로 종결 코돈)으로 번역하도록 지시한다. 일부 메테인생성 원핵생물에서 UAG 코돈(일반적으로 종결 코돈)은 피롤리신으로 번역될 수도 있다.[2]
진핵생물에는 표준 유전 부호의 20가지 아미노산에 셀레노시스테인을 더한 21가지의 단백질생성성 아미노산이 있다. 사람은 이들 단백질생성성 아미노산들 중 12가지를 서로 간 또는 다른 대사 중간생성물로부터 합성할 수 있다. 나머지 9가지는 음식물로부터 섭취해야 하기 때문에(보통 단백질 유도체로) 필수 아미노산이라고 한다. 필수 아미노산은 히스티딘, 아이소류신, 류신, 리신, 메티오닌, 페닐알라닌, 트레오닌, 트립토판, 발린이다.[3]
단백질생성성 아미노산은 리보자임의 자동아미노아실화 시스템에 의해 인식될 수 있는 아미노산의 세트와 관련이 있는 것으로 밝혀졌다.[4] 따라서 단백질비생성성 아미노산은 뉴클레오타이드 기반 생명체의 우발적인 진화적 성공에 의해 제외되었을 것이다. 특정 단백질비생성성 아미노산이 일반적으로 단백질에 통합되지 않는 이유를 설명하기 위해 다른 이유들이 제시되었다. 예를 들어 오르니틴과 호모세린은 펩타이드 골격에 대해 고리를 형성하고 상대적으로 짧은 반감기로 단백질을 파편화하는 반면, 다른 것들은 아르기닌 유사체인 카나바닌과 같이 단백질에 실수로 통합될 수 있기 때문에 독성이 있다.
원시 수프에서 특정 단백질생성성 아미노산의 진화적 선택은 단백질비생성성 아미노산과 달리 폴리펩타이드 사슬에 더 잘 통합되기 때문인 것으로 제안되었다.[5]
구조
[편집]다음 그림은 진핵생물의 유전 부호에 의해 단백질 합성에 직접적으로 사용되는 21가지 아미노산들의 구조와 약어이다. 그림에 나타난 구조는 수용액에서 존재하는 일반적인 양쪽성 이온의 형태가 아닌 표준 화학 구조이다.
 |
-
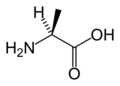 L-알라닌
L-알라닌
(Ala / A) -
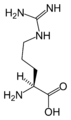 L-아르기닌
L-아르기닌
(Arg / R) -
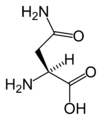 L-아스파라긴
L-아스파라긴
(Asn / N) -
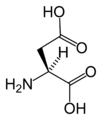 L-아스파르트산
L-아스파르트산
(Asp / D) -
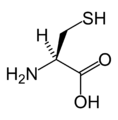 L-시스테인
L-시스테인
(Cys / C) -
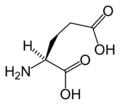 L-글루탐산
L-글루탐산
(Glu / E) -
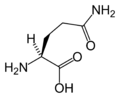 L-글루타민
L-글루타민
(Gln / Q) -
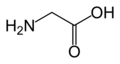 글리신
글리신
(Gly / G) -
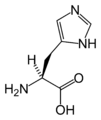 L-히스티딘
L-히스티딘
(His / H) -
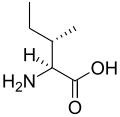 L-아이소류신
L-아이소류신
(Ile / I) -
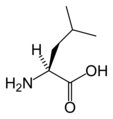 L-류신
L-류신
(Leu / L) -
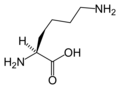 L-리신
L-리신
(Lys / K) -
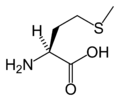 L-메티오닌
L-메티오닌
(Met / M) -
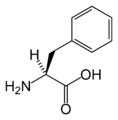 L-페닐알라닌
L-페닐알라닌
(Phe / F) -
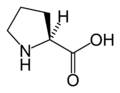 L-프롤린
L-프롤린
(Pro / P) -
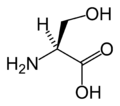 L-세린
L-세린
(Ser / S) -
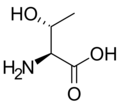 L-트레오닌
L-트레오닌
(Thr / T) -
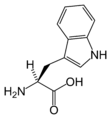 L-트립토판
L-트립토판
(Trp / W) -
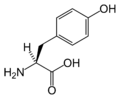 L-티로신
L-티로신
(Tyr / Y) -
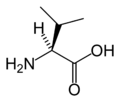 L-발린
L-발린
(Val / V)

.png)


















IUPAC 및 IUBMB는 현재 다음의 두 가지 아미노산에 대한 표준 약어도 권장한다.

화학적 특성
[편집]다음은 표준 아미노산의 한 문자 기호, 세 문자 기호 및 곁사슬의 화학적 특성을 나열한 표이다. 표시된 질량은 자연 존재비에서 동위원소의 가중 평균을 기반으로 한다. 펩타이드 결합을 형성하면 물 분자가 제거된다. 따라서 단백질의 질량은 펩타이드 결합당 18.01524 Da을 뺀 값으로 구성된 아미노산들의 질량과 같다.
일반 화학적 특성
[편집]| 아미노산 | 한 문자 약어 | 세 문자 약어 | 평균 질량 (Da) | pI | pK1 (α-COOH) |
pK2 (α-NH3+) |
|---|---|---|---|---|---|---|
| 알라닌 | A | Ala | 89.09404 | 6.01 | 2.35 | 9.87 |
| 시스테인 | C | Cys | 121.15404 | 5.05 | 1.92 | 10.70 |
| 아스파르트산 | D | Asp | 133.10384 | 2.85 | 1.99 | 9.90 |
| 글루탐산 | E | Glu | 147.13074 | 3.15 | 2.10 | 9.47 |
| 페닐알라닌 | F | Phe | 165.19184 | 5.49 | 2.20 | 9.31 |
| 글리신 | G | Gly | 75.06714 | 6.06 | 2.35 | 9.78 |
| 히스티딘 | H | His | 155.15634 | 7.60 | 1.80 | 9.33 |
| 아이소류신 | I | Ile | 131.17464 | 6.05 | 2.32 | 9.76 |
| 리신 | K | Lys | 146.18934 | 9.60 | 2.16 | 9.06 |
| 류신 | L | Leu | 131.17464 | 6.01 | 2.33 | 9.74 |
| 메티오닌 | M | Met | 149.20784 | 5.74 | 2.13 | 9.28 |
| 아스파라긴 | N | Asn | 132.11904 | 5.41 | 2.14 | 8.72 |
| 피롤리신 | O | Pyl | 255.31 | ? | ? | ? |
| 프롤린 | P | Pro | 115.13194 | 6.30 | 1.95 | 10.64 |
| 글루타민 | Q | Gln | 146.14594 | 5.65 | 2.17 | 9.13 |
| 아르기닌 | R | Arg | 174.20274 | 10.76 | 1.82 | 8.99 |
| 세린 | S | Ser | 105.09344 | 5.68 | 2.19 | 9.21 |
| 트레오닌 | T | Thr | 119.12034 | 5.60 | 2.09 | 9.10 |
| 셀레노시스테인 | U | Sec | 168.053 | 5.47 | 1.91 | 10 |
| 발린 | V | Val | 117.14784 | 6.00 | 2.39 | 9.74 |
| 트립토판 | W | Trp | 204.22844 | 5.89 | 2.46 | 9.41 |
| 티로신 | Y | Tyr | 181.19124 | 5.64 | 2.20 | 9.21 |
곁사슬의 특성
[편집]| 아미노산 | 한 문자 약어 | 세 문자 약어 | 곁사슬 | 소수성 | pKa§ | 극성 | pH | 작음 | 아주 작음 | 방향족 또는 지방족 |
반데르발스 부피 (Å3) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 알라닌 | A | Ala | -CH3 | - | - | 지방족 | 67 | ||||
| 시스테인 | C | Cys | -CH2SH | 8.55 | 산성 | - | 86 | ||||
| 아스파르트산 | D | Asp | -CH2COOH | 3.67 | 산성 | - | 91 | ||||
| 글루탐산 | E | Glu | -CH2CH2COOH | 4.25 | 산성 | - | 109 | ||||
| 페닐알라닌 | F | Phe | -CH2C6H5 | - | - | 방향족 | 135 | ||||
| 글리신 | G | Gly | -H | - | - | - | 48 | ||||
| 히스티딘 | H | His | -CH2-C3H3N2 | 6.54 | 약염기성 | 방향족 | 118 | ||||
| 아이소류신 | I | Ile | -CH(CH3)CH2CH3 | - | - | 지방족 | 124 | ||||
| 리신 | K | Lys | -(CH2)4NH2 | 10.40 | 염기성 | - | 135 | ||||
| 류신 | L | Leu | -CH2CH(CH3)2 | - | - | 지방족 | 124 | ||||
| 메티오닌 | M | Met | -CH2CH2SCH3 | - | - | 지방족 | 124 | ||||
| 아스파라긴 | N | Asn | -CH2CONH2 | - | - | - | 96 | ||||
| 피롤리신 | O | Pyl | -(CH2)4NHCOC4H5NCH3 | N.D. | 약염기성 | - | ? | ||||
| 프롤린 | P | Pro | -CH2CH2CH2- | - | - | - | 90 | ||||
| 글루타민 | Q | Gln | -CH2CH2CONH2 | - | - | - | 114 | ||||
| 아르기닌 | R | Arg | -(CH2)3NH-C(NH)NH2 | 12.3 | 강염기성 | - | 148 | ||||
| 세린 | S | Ser | -CH2OH | - | - | - | 73 | ||||
| 트레오닌 | T | Thr | -CH(OH)CH3 | - | - | - | 93 | ||||
| 셀레노시스테인 | U | Sec | -CH2SeH | 5.43 | 산성 | - | ? | ||||
| 발린 | V | Val | -CH(CH3)2 | - | - | 지방족 | 105 | ||||
| 트립토판 | W | Trp | -CH2C8H6N | - | - | 방향족 | 163 | ||||
| 티로신 | Y | Tyr | -CH2-C6H4OH | 9.84 | 약산성 | 방향족 | 141 |
§: 아스파르트산(Asp), 시스테인(Cys), 글루탐산(Glu), 히스티딘(His), 리신(Lys), 티로신(Tyr)에 대한 값은 알라닌 펜타펩타이드의 중앙에 배치된 아미노산 잔기를 사용하여 결정되었다.[6] 아르기닌(Arg)에 대한 값은 페이스(Pace) 등의 저자들의 논문(2009)[7]을 참조하였다. 셀레노시스테인(Sec)에 대한 값은 변(Byun)과 강(Kang)의 논문(2011)[8]을 참조하였다.
N.D.: 피롤리신의 pKa값은 보고되지 않았다.
주: 작은 펩타이드에 있는 아미노산 잔기의 pKa값은 일반저으로 단백질 내부에 있을 때와 약간 다르다. 이러한 상황에서 아미노산 잔기의 pKa값의 변화를 계산하기 위해 단백질 pKa 계산이 때때로 사용된다.
유전자 발현 및 생화학
[편집]| 아미노산 | 한 문자 약어 | 세 문자 약어 | 코돈 | 존재 | 사람에서 필수 아미노산‡ | |||
|---|---|---|---|---|---|---|---|---|
| 고세균의 단백질에 존재 (%)& |
세균의 단백질에 존재 (%)& |
진핵생물의 단백질에 존재 (%)& |
사람의 단백질에 존재 (%)& | |||||
| 알라닌 | A | Ala | GCU, GCC, GCA, GCG | 8.2 | 10.06 | 7.63 | 7.01 | 아니요 |
| 시스테인 | C | Cys | UGU, UGC | 0.98 | 0.94 | 1.76 | 2.3 | 조건부 |
| 아스파르트산 | D | Asp | GAU, GAC | 6.21 | 5.59 | 5.4 | 4.73 | 아니요 |
| 글루탐산 | E | Glu | GAA, GAG | 7.69 | 6.15 | 6.42 | 7.09 | 조건부 |
| 페닐알라닌 | F | Phe | UUU, UUC | 3.86 | 3.89 | 3.87 | 3.65 | 예 |
| 글리신 | G | Gly | GGU, GGC, GGA, GGG | 7.58 | 7.76 | 6.33 | 6.58 | 조건부 |
| 히스티딘 | H | His | CAU, CAC | 1.77 | 2.06 | 2.44 | 2.63 | 예 |
| 아이소류신 | I | Ile | AUU, AUC, AUA | 7.03 | 5.89 | 5.1 | 4.33 | 예 |
| 리신 | K | Lys | AAA, AAG | 5.27 | 4.68 | 5.64 | 5.72 | 예 |
| 류신 | L | Leu | UUA, UUG, CUU, CUC, CUA, CUG | 9.31 | 10.09 | 9.29 | 9.97 | 예 |
| 메티오닌 | M | Met | AUG | 2.35 | 2.38 | 2.25 | 2.13 | 예 |
| 아스파라긴 | N | Asn | AAU, AAC | 3.68 | 3.58 | 4.28 | 3.58 | 아니요 |
| 피롤리신 | O | Pyl | UAG* | 0 | 0 | 0 | 0 | 아니요 |
| 프롤린 | P | Pro | CCU, CCC, CCA, CCG | 4.26 | 4.61 | 5.41 | 6.31 | 아니요 |
| 글루타민 | Q | Gln | CAA, CAG | 2.38 | 3.58 | 4.21 | 4.77 | 아니요 |
| 아르기닌 | R | Arg | CGU, CGC, CGA, CGG, AGA, AGG | 5.51 | 5.88 | 5.71 | 5.64 | 조건부 |
| 세린 | S | Ser | UCU, UCC, UCA, UCG, AGU, AGC | 6.17 | 5.85 | 8.34 | 8.33 | 아니요 |
| 트레오닌 | T | Thr | ACU, ACC, ACA, ACG | 5.44 | 5.52 | 5.56 | 5.36 | 예 |
| 셀레노시스테인 | U | Sec | UGA** | 0 | 0 | 0 | >0 | 아니요 |
| 발린 | V | Val | GUU, GUC, GUA, GUG | 7.8 | 7.27 | 6.2 | 5.96 | 예 |
| 트립토판 | W | Trp | UGG | 1.03 | 1.27 | 1.24 | 1.22 | 예 |
| 티로신 | Y | Tyr | UAU, UAC | 3.35 | 2.94 | 2.87 | 2.66 | 조건부 |
| 종결 코돈† | - | Term | UAA, UAG, UGA†† | ? | ? | ? | 빈칸 | 빈칸 |
* UAG는 일반적으로 앰버 종결 코돈이지만 pylTSBCD 유전자 클러스터에 의해 암호화되는 생물학적 시스템을 포함한 생물에서는 피롤리신을 지정한다.[9]
** UGA는 일반적으로 오팔(또는 엄버) 종결 코돈이지만 SECIS 요소가 있는 경우 셀레노시스테인을 지정한다.
† 종결 코돈은 아미노산을 지정하지는 않지만 완전성을 위해 포함시킨다.
†† UAG 및 UGA가 항상 종결 코돈으로 작용하는 것은 아니다(위 참조).
‡ 필수 아미노산은 인체에서 합성할 수 없기 때문에 반드시 음식물로 섭취해야 한다. 조건부 필수 아미노산은 일반적으로 식단에 필요하지 않지만 적절한 양으로 합성되지 않는 특정 개체군에게는 외부로부터 공급되어야 한다.
& 아미노산의 존재 여부는 각각 135가지의 고세균 프로테옴, 3,775가지의 세균 프로테옴, 614가지의 진핵생물 프로테옴 및 사람의 프로테옴(21,006가지의 단백질)을 기반으로 한 것이다.[10]
질량분석법
[편집]펩타이드와 단백질의 질량분석법에서 잔기의 질량에 대한 지식은 유용하다. 펩타이드 또는 단백질의 질량은 잔기의 질량과 물의 질량의 합(단일 동위원소 질량 = 18.01056 Da, 평균 질량 = 18.0153 Da)이다. 잔기의 질량은 화학식 및 원자량으로부터 계산된다.[11] 질량분석법에서 이온은 하나 이상의 양성자를 포함할 수도 있다(단일 동위원소 질량 = 1.00728 Da, 평균 질량* = 1.0074 Da). *양성자는 평균 질량을 가질 수 없다. 이것은 중양성자를 유효한 동위원소로 혼란스럽게 추론하지만, 이들은 다른 화학종이어야 한다(히드론을 참조).
| 아미노산 | 한 문자 약어 | 세 문자 약어 | 화학식 | 단일 동위원소 질량§ (Da) | Avg. mass (Da) |
|---|---|---|---|---|---|
| 알라닌 | A | Ala | C3H5NO | 71.03711 | 71.0779 |
| 시스테인 | C | Cys | C3H5NOS | 103.00919 | 103.1429 |
| 아스파르트산 | D | Asp | C4H5NO3 | 115.02694 | 115.0874 |
| 글루탐산 | E | Glu | C5H7NO3 | 129.04259 | 129.1140 |
| 페닐알라닌 | F | Phe | C9H9NO | 147.06841 | 147.1739 |
| 글리신 | G | Gly | C2H3NO | 57.02146 | 57.0513 |
| 히스티딘 | H | His | C6H7N3O | 137.05891 | 137.1393 |
| 아이소류신 | I | Ile | C6H11NO | 113.08406 | 113.1576 |
| 리신 | K | Lys | C6H12N2O | 128.09496 | 128.1723 |
| 류신 | L | Leu | C6H11NO | 113.08406 | 113.1576 |
| 메티오닌 | M | Met | C5H9NOS | 131.04049 | 131.1961 |
| 아스파라긴 | N | Asn | C4H6N2O2 | 114.04293 | 114.1026 |
| 피롤리신 | O | Pyl | C12H19N3O2 | 237.14773 | 237.2982 |
| 프롤린 | P | Pro | C5H7NO | 97.05276 | 97.1152 |
| 글루타민 | Q | Gln | C5H8N2O2 | 128.05858 | 128.1292 |
| 아르기닌 | R | Arg | C6H12N4O | 156.10111 | 156.1857 |
| 세린 | S | Ser | C3H5NO2 | 87.03203 | 87.0773 |
| 트레오닌 | T | Thr | C4H7NO2 | 101.04768 | 101.1039 |
| 셀레노시스테인 | U | Sec | C3H5NOSe | 150.95364 | 150.0489 |
| 발린 | V | Val | C5H9NO | 99.06841 | 99.1311 |
| 트립토판 | W | Trp | C11H10N2O | 186.07931 | 186.2099 |
| 티로신 | Y | Tyr | C9H9NO2 | 163.06333 | 163.1733 |
세포의 화학량론 및 대사 비용
[편집]아래의 표는 대장균 세포에서의 풍부한 아미노산과 아미노산 합성을 위한 대사 비용(ATP)을 나열한 것이다. 음수는 대사 과정이 에너지적으로 유리하고 세포의 순ATP를 소모하지 않음을 나타낸다.[12] 풍부한 아미노산은 유리된 형태와 중합된 형태(단백질)의 아미노산을 포함한 것이다.
| 아미노산 | 한 문자 약어 | 세 문자 약어 | 풍부도 (대장균 세포당of molecules 분자수 (×108)) |
합성시 ATP 비용 | |
|---|---|---|---|---|---|
| 호기성 조건 |
혐기성 조건 | ||||
| 알라닌 | A | Ala | 2.9 | -1 | 1 |
| 시스테인 | C | Cys | 0.52 | 11 | 15 |
| 아스파르트산 | D | Asp | 1.4 | 0 | 2 |
| 글루탐산 | E | Glu | 1.5 | -7 | -1 |
| 페닐알라닌 | F | Phe | 1.1 | -6 | 2 |
| 글리신 | G | Gly | 3.5 | -2 | 2 |
| 히스티딘 | H | His | 0.54 | 1 | 7 |
| 아이소류신 | I | Ile | 1.7 | 7 | 11 |
| 리신 | K | Lys | 2.0 | 5 | 9 |
| 류신 | L | Leu | 2.6 | -9 | 1 |
| 메티오닌 | M | Met | 0.88 | 21 | 23 |
| 아스파라긴 | N | Asn | 1.4 | 3 | 5 |
| 피롤리신 | O | Pyl | - | - | - |
| 프롤린 | P | Pro | 1.3 | -2 | 4 |
| 글루타민 | Q | Gln | 1.5 | -6 | 0 |
| 아르기닌 | R | Arg | 1.7 | 5 | 13 |
| 세린 | S | Ser | 1.2 | -2 | 2 |
| 트레오닌 | T | Thr | 1.5 | 6 | 8 |
| 셀레노시스테인 | U | Sec | - | - | - |
| 발린 | V | Val | 2.4 | -2 | 2 |
| 트립토판 | W | Trp | 0.33 | -7 | 7 |
| 티로신 | Y | Tyr | 0.79 | -8 | 2 |
비고
[편집]| 아미노산 | 약어 | 비고 | |
|---|---|---|---|
| 알라닌 | A | Ala | 알라닌은 매우 풍부하고 다재다능하며 글리신보다 더 뻣뻣하지만 단백질의 입체형태에 대해 작은 입체적 한계를 나타낼 만큼 충분히 작다. 알라닌은 상당히 중립적으로 행동하며 단백질 외부의 친수성 영역과 내부의 소수성 영역 모두에 위치할 수 있다. |
| 아스파라긴 또는 아스파르트산 | B | Asx | 아미노산 중 하나가 위치를 차지할 수 있는 자리 표시자. |
| 시스테인 | C | Cys | 황 원자는 중금속 이온에 쉽게 결합한다. 산화되는 조건에서 두 개의 시스테인이 이황화 결합으로 결합하여 시스틴을 형성할 수 있다. 시스틴이 단백질의 일부인 경우(예: 인슐린) 3차 구조가 안정화되어 단백질이 변성에 더 잘 견디게 된다. 따라서 이황화 결합은 소화 효소(예: 펩신 및 키모트립신) 및 구조 단백질(예: 케라틴)을 포함한 가혹한 환경에서 기능을 해야 하는 단백질에서 일반적으로 발견된다. 이황화물은 자체적으로 안정적인 모양을 유지하기에는 너무 작은 펩타이드(예: 인슐린)에서도 발견된다. |
| 아스파르트산 | D | Asp | 아스파르트산은 글루탐산과 유사하게 행동하며 강한 음전하를 갖는 친수성 산성기를 운반한다. 아스파르트산은 일반적으로 단백질의 외부 표면에 위치하여 단백질을 수용성으로 만든다. 아스파르트산은 양전하를 띤 분자와 이온에 결합하여 보통 효소에서 금속 이온을 고정시키는 데 사용된다. 단백질 내부에 위치할 때, 아스파르트산 및 글루탐산은 일반적으로 아르기닌 및 리신과 짝을 이룬다. |
| 글루탐산 | E | Glu | 글루탐산은 아스파르트산과 유사하게 행동하며 더 길고 약간 더 유연한 곁사슬을 가지고 있다. |
| 페닐알라닌 | F | Phe | 사람에게 필수적인 페닐알라닌, 티로신, 트립토판은 곁사슬에 크고 단단한 방향족 작용기를 포함하고 있다. 이들은 크기가 큰 아미노산들이다. 아이소류신, 류신, 발린과 마찬가지로 이들 아미노산은 소수성이며 접힌 단백질에서 분자의 내부를 향하는 경향이 있다. 페닐알라닌은 티로신으로 전환될 수 있다. |
| 글리신 | G | Gly | α-탄소에 있는 두 개의 수소 원자 때문에 글리신은 광학적으로 활성이 아니다. 글리신은 가장 작은 아미노산으로 쉽게 회전하며 단백질 사슬에 유연성을 부여한다. 글리신은 콜라겐의 삼중 나선과 같은 가장 좁은 공간에 들어갈 수 있다. 너무 많은 유연성은 일반적으로 바람직하지 않기 때문에 구조적 구성 요소로서 글리신은 알라닌보다 덜 일반적이다. |
| 히스티딘 | H | His | 히스티딘은 사람에게 필수적이다. 약간 산성인 조건에서도 질소의 양성자화가 일어나 히스티딘과 폴리펩타이드의 특성이 전체적으로 변한다. 이것은 조절 메커니즘으로 많은 단백질에 의해 사용되며, 후기 엔도솜 또는 리소좀과 같은 산성 영역에서 폴리펩타이드의 형태와 행동을 변화시켜 효소의 입체형태의 변화를 강화한다. 그러나 이를 위해서는 소수의 히스티딘만 필요하기 때문에 히스타민은 비교적 희소하다. |
| 아이소류신 | I | Ile | 아이소류신은 사람에게 필수적이다. 아이소류신, 류신, 발린은 큰 지방족 소수성 곁사슬을 가지고 있다. 이들 분자는 단단하고, 이들 사슬은 단백질 분자 내부에 위치하는 경향이 있기 때문에 이들 상호간의 소수성 상호작용은 단백질의 올바른 접힘에 중요하다. |
| 류신 또는 아이소류신 | J | Xle | 아미노산 중 하나가 위치를 차지할 수 있는 자리 표시자. |
| 리신 | K | Lys | 리신은 사람에게 필수적이며 아르기닌과 유사하게 행동한다. 리신은 양전하를 띤 길고 유연한 곁사슬을 가지고 있다. 사슬의 유연성으로 인해 리신과 아르기닌은 표면에 음전하가 많은 분자와 결합하기에 적합하다. 예를 들어 DNA 결합 단백질은 아르기닌과 리신이 풍부한 활성 영역을 가지고 있다. 강한 전하로 인해 이 두 아미노산은 단백질의 친수성 외부 표면에 쉽게 위치하게 된다. 내부에서 발견되면 일반적으로 음전하를 띤 아미노산(예: 아스파르트산 또는 글루탐산)과 짝을 이룬다. |
| 류신 | L | Leu | 류신은 사람에게 필수적이며 아이소류신 및 발린과 유사하게 행동한다. |
| 메티오닌 | M | Met | 메티오닌은 사람에게 필수적이다. 단백질에 통합되는 첫 번째 아미노산이며, 번역 후에 제거되기도 한다. 시스테인과 마찬가지로 황을 포함하고 있지만 수소 대신에 메틸기가 있다. 이 메틸기는 활성화될 수 있으며, 새로운 탄소 원자가 다른 분자에 첨가되는 많은 반응에 사용된다. |
| 아스파라긴 | N | Asn | 카복실기를 가지고 있는 아스파르트산과 유사하며, 아스파라긴은 아마이드기를 가지고 있다. |
| 피롤리신 | O | Pyl | 피롤리신은 리신과 유사하지만 피롤린 고리가 붙어 있다. |
| 프롤린 | P | Pro | 프롤린은 N-말단의 아미노기에 특이한 고리를 포함하고 있으며 CO-NH 아마이드 서열을 고정된 입체형태로 만든다. 이것은 α 나선 또는 β 시트와 같은 단백질 접힘 구조를 파괴하여 단백질 사슬에 원하는 꼬임을 강요할 수 있다. 콜라겐에서 흔히 볼 수 있으며, 종종 번역 후 변형에 의해 하이드록시프롤린으로 변형된다. |
| 글루타민 | Q | Gln | 카복실기를 가지고 있는 글루탐산과 유사하며, 글루타민은 아마이드기를 가지고 있다. 글루타민은 단백질에서 사용되며 암모니아를 저장하는 데 사용되고, 몸에서 가장 풍부한 아미노산이다. |
| 아르기닌 | R | Arg | 기능적으로 리신과 유사하다. |
| 세린 | S | Ser | 세린과 트레오닌은 하이드록실기로 끝나는 짧은 곁사슬을 가지고 있다. 세린과 트레오닌의 수소는 제거하기 쉽기 때문에 세린과 트레오닌은 종종 효소에서 수소 공여체로 작용한다. 둘 다 매우 친수성이기 때문에 용해성 단백질의 외부 영역에는 세린과 트레오닌이 풍부한 경향이 있다. |
| 트레오닌 | T | Thr | 트레오닌은 사람에게 필수적이며 세린과 유사하게 행동한다. |
| 셀레노시스테인 | U | Sec | 황 원자가 셀레늄으로 치환된 시스테인의 셀레늄 유사체이다. |
| 발린 | V | Val | 발린은 사람에게 필수적이며 아이소류신 및 류신과 유사하게 행동한다. |
| 트립토판 | W | Trp | 트립토판은 사람에게 필수적이며 페닐알라닌 및 티로신과 유사하게 행동한다. 트립토판은 세로토닌의 전구체이며 자연적으로 형광성이다. |
| 알려지지 않은 | X | Xaa | 아미노산이 알려지지 않았거나 중요하지 않은 경우의 자리 표시자. |
| 티로신 | Y | Tyr | 티로신은 페닐알라닌(티로신의 전구체) 및 트립토판과 유사하게 행동하며, 멜라닌, 에피네프린, 갑상샘 호르몬의 전구체이다. 자연적으로 형광성이며 일반적으로 트립토판으로의 에너지 전달에 의해 형광성이 소멸된다. |
| 글루탐산 또는 글루타민 | Z | Glx | 아미노산 중 하나가 위치를 차지할 수 있는 자리 표시자. |
이화작용
[편집] |
아미노산은 주요 생성물의 특성에 따라 다음과 같이 분류할 수 있다.[13]
- 포도당생성성 아미노산 – 포도당신생합성에 의해 포도당으로 전환될 수 있는 아미노산이다.
- 케톤체생성성 아미노산 – 케톤체의 전구체인 아세틸-CoA로 직접적으로 분해될 수 있는 아미노산이다. 이들은 케톤체생성 또는 지질 합성에 사용될 수 있다.
- 포도당생성성 아미노산이면서 케톤체생성성 아미노산이기도 한 아미노산
같이 보기
[편집]각주
[편집]- ↑ Ambrogelly A, Palioura S, Söll D (January 2007). “Natural expansion of the genetic code”. 《Nature Chemical Biology》 3 (1): 29–35. doi:10.1038/nchembio847. PMID 17173027.
- ↑ Lobanov AV, Turanov AA, Hatfield DL, Gladyshev VN (August 2010). “Dual functions of codons in the genetic code”. 《Critical Reviews in Biochemistry and Molecular Biology》 45 (4): 257–65. doi:10.3109/10409231003786094. PMC 3311535. PMID 20446809.
- ↑ Young VR (August 1994). “Adult amino acid requirements: the case for a major revision in current recommendations” (PDF). 《The Journal of Nutrition》 124 (8 Suppl): 1517S–1523S. doi:10.1093/jn/124.suppl_8.1517S. PMID 8064412.
- ↑ Erives A (August 2011). “A model of proto-anti-codon RNA enzymes requiring L-amino acid homochirality”. 《Journal of Molecular Evolution》 73 (1–2): 10–22. Bibcode:2011JMolE..73...10E. doi:10.1007/s00239-011-9453-4. PMC 3223571. PMID 21779963.
- ↑ Frenkel-Pinter, Moran; Haynes, Jay W.; C, Martin; Petrov, Anton S.; Burcar, Bradley T.; Krishnamurthy, Ramanarayanan; Hud, Nicholas V.; Leman, Luke J.; Williams, Loren Dean (2019년 8월 13일). “Selective incorporation of proteinaceous over nonproteinaceous cationic amino acids in model prebiotic oligomerization reactions”. 《Proceedings of the National Academy of Sciences》 (영어) 116 (33): 16338–16346. doi:10.1073/pnas.1904849116. ISSN 0027-8424. PMC 6697887. PMID 31358633.
- ↑ Thurlkill RL, Grimsley GR, Scholtz JM, Pace CN (May 2006). “pK values of the ionizable groups of proteins”. 《Protein Science》 15 (5): 1214–8. doi:10.1110/ps.051840806. PMC 2242523. PMID 16597822.
- ↑ Pace CN, Grimsley GR, Scholtz JM (May 2009). “Protein ionizable groups: pK values and their contribution to protein stability and solubility”. 《The Journal of Biological Chemistry》 284 (20): 13285–9. doi:10.1074/jbc.R800080200. PMC 2679426. PMID 19164280.
- ↑ Byun BJ, Kang YK (May 2011). “Conformational preferences and pK(a) value of selenocysteine residue”. 《Biopolymers》 95 (5): 345–53. doi:10.1002/bip.21581. PMID 21213257. S2CID 11002236.
- ↑ Rother M, Krzycki JA (August 2010). “Selenocysteine, pyrrolysine, and the unique energy metabolism of methanogenic archaea”. 《Archaea》 2010: 1–14. doi:10.1155/2010/453642. PMC 2933860. PMID 20847933.
- ↑ Kozlowski LP (January 2017). “Proteome-pI: proteome isoelectric point database”. 《Nucleic Acids Research》 45 (D1): D1112–D1116. doi:10.1093/nar/gkw978. PMC 5210655. PMID 27789699.
- ↑ “Atomic Weights and Isotopic Compositions for All Elements”. NIST. 2016년 12월 12일에 확인함.
- ↑ Phillips R, Kondev J, Theriot J, Garcia HG, Orme N (2013). 《Physical biology of the cell》 Seco판. Garland Science. 178쪽. ISBN 978-0-8153-4450-6.
- ↑ Ferrier DR (2005). 〈Chapter 20: Amino Acid Degradation and Synthesis〉. Champe PC, Harvey RA, Ferrier DR. 《Lippincott's Illustrated Reviews: Biochemistry (Lippincott's Illustrated Reviews)》. Hagerstwon, MD: Lippincott Williams & Wilkins. ISBN 978-0-7817-2265-0.
참조 문헌
[편집]- Nelson, David L.; Cox, Michael M. (2000). 《Lehninger Principles of Biochemistry》 3판. Worth Publishers. ISBN 978-1-57259-153-0.
- Kyte J, Doolittle RF (May 1982). “A simple method for displaying the hydropathic character of a protein”. 《Journal of Molecular Biology》 157 (1): 105–32. CiteSeerX 10.1.1.458.454. doi:10.1016/0022-2836(82)90515-0. PMID 7108955.
- Meierhenrich, Uwe J. (2008). 《Amino acids and the asymmetry of life》 1판. Springer. ISBN 978-3-540-76885-2.
- Biochemistry, Harpers (2015). 《Harpers Illustrated Biochemistry》 30판. Lange. ISBN 978-0-07-182534-4.
