Tutorial 29, Appendix B: good, bad, and ugly titles of Matt’s papers
February 26, 2015
Last October, Mike posted a tutorial on how to choose a paper title, then followed it up by evaluating the titles of his own papers. He invited me to do the same for my papers. I waited a few days to allow myself to forget Mike’s comments on our joint papers – not too hard during my fall anatomy teaching – and then wrote down my thoughts.
And then did nothing with them for three and a half months.
The other day I rediscovered that draft and thought, hey, I don’t remember anything I wrote back then, I should redo the experiment and see if my evaluations will be consistent. And this time without looking at Mike’s post at all, so the risk of contamination would be even lower.
BUT FIRST I thought I should write down what I admire in paper titles, so I could see whether my titles actually lived up to my ideals. So now we can compare:
- what I say I like in paper titles;
- what I actually titled my papers;
- what I had to say about my titles last October;
- what I have to say about them now;
- and, for some of my papers, what Mike had to say about them.
What I Admire In Paper Titles
Brevity. I first became consciously aware of the value of concise titles when I read Knut Schmidt-Nielsen’s autobiography, The Camel’s Nose, in 2004 or 2005. (Short-short review: most of the book is a narrative about scientific questions and it’s great, the self-congratulatory chapters near the end are much less interesting. Totally worth reading, especially since used copies can be had for next to nothing.) Schmidt-Nielsen said he always preferred short, simple titles. Short titles are usually punchy and hard to misunderstand. And I like titles that people can remember, and a short title is easier to recall than a long one.
Impact. In short, maximum information transfer using the minimum number of words. This is a separate point from sheer brevity; a paper can have a short title that doesn’t actually tell you very much. But brevity helps, because it’s difficult to compose a long title that really hits hard. Whatever impact a title might have, it will be diluted by every extraneous word.
Full sentences as titles. This is taking the information-transfer aspect of the last admirable quality to its logical extreme, although often at the expense of brevity. I was heavily influenced here by two things that happened while I was at Berkeley. First, I taught for a year in an NSF GK-12 program, where graduate students went out into local elementary, middle, and high schools and taught biology enrichment classes. One thing that was drilled into us during that experience is that we were teaching concepts, which ideally would be expressed as complete sentences. Also about that same time I read James Valentine’s book On the Origin of Phyla. The table of contents of that book is several pages long, because every chapter title, heading, and subheading is a complete sentence. This has a lovely effect: once you’ve read the table of contents of the book or any of its parts, you’ve gotten the TL;DR version of the argument. Sort of like a distributed abstract. I’d like to do that more.
How Did I Do?
Time to see if my actions match my words. Full bibliographic details and PDFs are available on my publications page. I stuck with Mike’s red-blue-green color scheme for the verdicts. My October 2014 and February 2015 thoughts are labeled. For joint papers with Mike, I’ve copied his assessment in as well. Any comments in brackets are my editorializing now, comparing what I said in October to what I said a few days ago before I’d looked back at my old comments or Mike’s.
* * * * * * * * * * * *
Sauroposeidon proteles, a new sauropod from the Early Cretaceous of Oklahoma. (11 words)
GOOD
Oct 2014: Like it. Short, to the point, includes the taxon name.
Feb 2015: Good, gets the job done with a minimum of fuss
Osteology, paleobiology, and relationships of the sauropod dinosaur Sauroposeidon. (9 words)
OK
Oct 2014: This title was inspired by the papers from the early 20th century
Feb 2015: It gets the job done, I suppose. I can’t help but wonder if there might have been a more elegant solution. Part of my unease is that this title is an example of the same attitude that produced the next monstrosity.
Osteological correlates of cervical musculature in Aves and Sauropoda (Dinosauria: Saurischia), with comments on the cervical ribs of Apatosaurus. (19 words)
BAD
Oct 2014: Ugh. It gets the job done, I suppose, but it’s waaaay long and just kind of ugly.
Feb 2015: Ugh. Waaay too wordy. I had a (fortunately brief) fascination with long titles, and especially the phrase, “with comments on”. Now I would cut it down to “Bony correlates of neck muscles in birds and sauropod dinosaurs” (10 words)
Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs. (10 words)
OK
Oct 2014: Like it. Would be better made into a sentence, like, “Vertebral pneumaticity is evidence for air sacs in sauropod dinosaurs.”
Feb 2015: Fairly clean. Does what it says on the tin. I’m having a hard time seeing how it could be turned into a sentence and still convey so much of what the paper is about in so few words.
[Heh. As we will see again later on, I was evidently smarter last fall than I am now.]
The evolution of vertebral pneumaticity in sauropod dinosaurs. (8 words)
GREAT
Oct 2014: Like it. It couldn’t really be any shorter without losing crucial information. Happy to have a decent title on my second-most-cited paper!
Feb 2015: Short, clean, probably my best title ever.
First occurrence of Brachiosaurus (Dinosauria: Sauropoda) from the Upper Jurassic Morrison Formation of Oklahoma. (14 words)
OK
Oct 2014: Yep. once you’ve read the title, you barely need to read the paper. Even better would have been, “A metacarpal of Brachiosaurus from the Upper Jurassic Morrison Formation of Oklahoma.” (12 words)
Feb 2015: Does what it says, but like my other PaleoBios pub, it’s a long title for a short paper. Now I would title it, “First record of the sauropod dinosaur Brachiosaurus from Oklahoma” (9 words)
[my October title was better!]
Postcranial skeletal pneumaticity in sauropods and its implications for mass estimates. (11 words)
OK
Oct 2014: It’s not elegant but it gets the job done. I wanted that paper to be one-stop shopping for sauropod PSP, but of course the real payoff there is the ASP/mass-estimate stuff, so I’m happy to have punched that up in the title.
Feb 2015: Good enough. I like it. It’s a little long–I could reasonably have just titled this, “Postcranial skeletal pneumaticity in sauropods”, but I wanted to draw attention to the implications for mass estimates.
Sauroposeidon: Oklahoma’s native giant (4 words)
Origin of postcranial skeletal pneumaticity in dinosaurs. (7 words)
GOOD
Oct 2014: About all I would change now would be to add the word “early” at the beginning of the title.
Feb 2015: Great. Could not be shortened further without losing information.
What pneumaticity tells us about ‘prosauropods’, and vice versa. (9 words)
GOOD
Oct 2014: Love this title. I used it for the abstract of the SVP talk that the paper was derived from, too.
Feb 2015: Kind of a gimmick title, but it’s accurate–the SVP abstract this paper was based on was built around a bullet list. And it’s still nice and short.
Evidence for bird-like air sacs in saurischian dinosaurs. (9 words)
GOOD
Oct 2014: Along with Wedel (2003b) and Wedel (2006), this has a short (7-9 words apiece) title that tells you what’s in the paper, simply and directly. For once, I’m glad I didn’t turn it into a sentence. I think a declarative statement like “Saurischian dinosaurs had air sacs like those of birds” would have been less informative and come off as advertising. I wanted this paper to do what the title said: run down the evidence for air sacs in saurischians.
Feb 2015: I like it and wouldn’t change it. The “evidence for” part is key – I didn’t want to write a paper primarily about the air sacs themselves. Instead I wanted to lay out the evidence explaining why we think sauropods had air sacs.
Head and neck posture in sauropod dinosaurs inferred from extant animals. (8 words)
OK
Oct 2014: It’s not horrible but it would be better as a declarative statement like, “Sauropod dinosaurs held their necks and heads elevated like most other tetrapods.” (12 words)
Feb 2015: Good. Reads almost telegraphically brief as it is. Does what it says on the tin.
Mike: RUBBISH
[October Matt wins again!]
A new sauropod dinosaur from the Lower Cretaceous Cedar Mountain Formation, Utah, USA. (13 words)
OK
Oct 2014: Two things about this one. First, I wish we’d been able to include the taxon name in the title, as we were allowed to do back in the day for Sauroposeidon. Second, I know some people whinge about us using the CMF in the title and in the paper instead of the Burro Canyon Fm, which is what the CMF is technically called east of the Colorado River. But srsly, how many people search for Burro Canyon Fm versus CMF? All of the relevant faunal comparisons are to be made with the CMF, so I don’t feel the least bit bad about this.
Feb 2015: Fine. About as short as it could be and still be informative.
Mike: RUBBISH
The long necks of sauropods did not evolve primarily through sexual selection. Journal of Zoology. (12 words)
GOOD
Oct 2014: Perfect. The abstract and the paper expand on the title, but if all you read is the title, you know what we found. That’s a worthy goal.
Feb 2015: My first sentence title. Every word does work, so even though this is one of my longer titles, I like it. The length relative to my other titles is not a knock against this one; rather, it emphasizes how well I did at keeping my early titles short and to the point (with a couple of regrettable exceptions as noted above).
Mike: SWEET
The early evolution of postcranial skeletal pneumaticity in sauropodomorph dinosaurs. (10 words)
GOOD
Oct 2014: Not bad. I wonder if something like, “Widespread vertebral fossae show that pulmonary pneumaticity evolved early in sauropodomorphs” might be better. It’s hard, though, to put so many long, polysyllabic words in a title that doesn’t sound like a train wreck. At a minimum, this paper does what it says on the tin.
Feb 2015: Short and to the point. Another one that couldn’t be any shorter without losing valuable information.
A monument of inefficiency: the presumed course of the recurrent laryngeal nerve in sauropod dinosaurs. (15 words)
Objectively: BAD to OK
Subjectively: GOOD to FREAKIN’ AWESOME
Oct 2014: I readily admit that I could have fashioned a more informative title, but I dearly love this one. It’s derived from a TV commercial for cheeseburgers (true story), and it warms my heart every time I read it.
Feb 2015: This is definitely a gimmick title that is longer than it has to be (it would be a concise 11 words without the unnecessary intro clause) BUT I love it and I’d do it exactly the same if I could do it again. So there!
Why sauropods had long necks; and why giraffes have short necks. (11 words)
GOOD
Oct 2014: This is one of those ‘draw the reader in’ titles. I like it.
Feb 2015: We both liked the even shorter, “Why giraffes have short necks” but we really felt that a paper about sauropod necks needed sauropod necks in the title. I feel about this one like I feel about my 2007 prosauropod paper: it’s a gimmick title, but it’s short, so no harm done.
Mike: EXCELLENT
Neural spine bifurcation in sauropod dinosaurs of the Morrison Formation: ontogenetic and phylogenetic implications. (14 words)
OK
Oct 2014: Blah. It’s okay, not great. Maybe better as, “No evidence for increasing neural spine bifurcation through ontogeny in diplodocid sauropods of the Morrison Formation”, or something along those lines.
Feb 2015: This one is long but I think the length is necessary. It’s also kinda boring, but it was addressing a fairly dry point. I think any attempt to shorten it or sexy it up would come off as gratuitous.
Mike: WEAK
The effect of intervertebral cartilage on neutral posture and range of motion in the necks of sauropod dinosaurs. (18 words)
OK
Oct 2014: Probably better along the lines of, “Intervertebral spacing suggests a high neutral posture and broad range of motion in the necks of sauropod dinosaurs” or something like that.
Feb 2015: My second-longest title ever! Looking at it now, I think we could have titled it, “Effects of intervertebral cartilage on neck posture and range of motion in sauropod dinosaurs” and gotten it down to 14 words, but the word ‘neutral’ is doing real work in the original so maybe that’s a bust.
Mike: UGH, rubbish.
[October Matt is up by three points at least]
Caudal pneumaticity and pneumatic hiatuses in the sauropod dinosaurs Giraffatitan and Apatosaurus. (12 words)
OK
Oct 2014: Along the same lines as the previous: “Caudal pneumaticity and pneumatic hiatuses show that pulmonary diverticula in the tails of sauropod dinosaurs were pervasive and complex” or something.
Feb 2015: Good. Long only by comparison with some of my earlier titles. Does what it says.
Mike: NOT GOOD ENOUGH
The neck of Barosaurus was not only longer but also wider than those of Diplodocus and other diplodocines. (18 words)
GOOD
Feb 2015: My second sentence-as-title, and another entry in the run of mostly long titles from 2012 onward. I like how precise it is, despite the length.
Mike: GOOD
A ceratopsian dinosaur from the Lower Cretaceous of Western North America, and the biogeography of Neoceratopsia. (16 words)
GOOD
Feb 2015: I had no say in this one (by choice, I’m sure Andy et al. would have listened if I had had any suggestions about the title, but I didn’t). If I could rewrite it, I’d probably make it even longer by adding in the word ‘new’ between A and ceratopsian
Haplocanthosaurus (Saurischia: Sauropoda) from the lower Morrison Formation (Upper Jurassic) near Snowmass, Colorado. (13 words)
OK
Feb 2015: Feels a lot longer than its 13 words, mostly because so many of the words are polysyllabic. Normally I like pulling the words in parentheses out, but in this case I can’t see that doing that would actually improve the title. Sometimes descriptive papers need plain titles. It’s okay.
* * * * * * * * * * * *
Lessons
First, Mike graded harder than I did. In fact, I only rated one of my titles as BAD, which seems a bit feeble. I think we were using different criteria. If a title was boring but serviceable, I gave it an OK, whereas Mike tended to flag any suboptimal title as RUBBISH. But I didn’t remember that about his post, and I deliberately avoided looking at it until I’d made my evaluations.
Second, except for the two PaleoBios papers, all of the titles from the first half of my career (2000-2007) are 12 words or fewer, including a substantial bundle from before I’d read either The Camel’s Nose or Strunk & White. I’m sure that being a Cifelli student and then a Padian student had something to do with that; Rich and Kevin made me into the word choice and grammar pedant that I am today (my rhetorical excrescences on this site are my fault, not theirs).
Third, much to my surprise and consternation, my titles have gotten longer over time, not shorter. Partly that’s because my little corner of the science ecosystem is getting increasingly subdivided, so it’s hard for me to write a paper now with a title as broad as, “The evolution of vertebral pneumaticity in sauropod dinosaurs.” (Possibly a prod to keep seeking out new, more open horizons?) And I suppose there is some tension between brevity, informativeness, and precision. For example, saying in the title of a descriptive paper than a specimen is “from the Upper Jurassic Morrison Formation of [Location], [State or Country]” adds 11 words, but the title really does need those words. That could be a segue into a whole other discussion about descriptive versus analytical work, but that will be a topic for another time.
Ultimately, this has been a fun exercise and it’s made me more aware of how I title my papers. This is useful because I have some manuscripts in the works that deal with really detailed anatomy, and I need to figure out how to give them titles that are precise and informative but still punchy. It’s not easy.
Parting thought: after I posted the slides from my photography and illustration talk, Mike and I talked about posting some of our figures and dissecting them to see how they could be improved (it’s axiomatic that almost all figures could be improved in one way or another). We should really get started on that.
Dinosaur National Monument quarry map
February 25, 2015
The Carnegie Quarry, at Dinosaur National Monument, near Jensen, Utah, is arguably the most impressive dinosaur-fossil exhibit anywhere in the world — a covered, semi-excavated quarry that’s absolutely packed with big dinosaur fossils.
It’s also notoriously difficult to photograph: too big to fit into a single photo, and with poor contrast between the bones and matrix. This is the best picture I’ve found of part of it (from here) …

… although this one (from here) conveys the scale better:

It’s one of the great sadnesses of my life that I’ve yet to visit DNM.
The quarry is historically important: discovered by Earl Douglass in 1909, it yielded among other specimens CM 3018, the holotype of Apatosaurus louisae and the principle subject of Gilmore’s (1936) monograph.
I’ve only recently become aware (thanks, Matt!) of Ken Carpenter’s (2013) detailed treatment of the history, sedimentology and taphonomy of the quarry — an important work that deserves to be widely read. Pages 10-14 are largely taken up with parts A-E of figure 10 — a big multi-page map of the quarry, showing the location of its most important specimens. Unfortunately, the five sections of this figure are all at slightly different scales in the PDF. I’ve rescaled them and pasted them together into a single big (4387 × 1210) image which I reproduce here:

Carpenter (2013:fig 10): map of the Carnegie quarry, composited from parts A-E.
Enjoy!
Update (six hours later)
I just heard from Ken Carpenter, who created the illustration. He has kindly sent me the full-resolution version — which is four times as big as the one I extracted from the PDF — and gave me permission to post it here on SV-POW! under the CC By licence. So here it is!

Thanks, Ken!
Second update (12 March 2015)
Over on the Extinct Monsters blog, Ben Miller has published The Carnegie Quarry Diaspora. It’s a beautiful illustrated survey of some of the most important specimens to have come out of this quarry, including no fewer than seven important sauropod individuals.
References
- Carpenter, Kenneth. 2013. History, sedimentology, and taphonomy of the Carnegie Quarry, Dinosaur National Monument, Utah. Annals of the Carnegie Museum 81(3):153-232.
- Gilmore, C. W. 1936. Osteology of Apatosaurus with special reference to specimens in the Carnegie Museum. Memoirs of the Carnegie Museum 11: 175-300.
Defensive use of the tail in monitors – and also sauropods?
February 22, 2015

Murphy and Mitchell (1974: fig. 1)
One thing that I’ve never understood is why some people are skeptical about sauropods using their tails defensively, when lizards do this all the time. I’ve been digging through the literature on this for a current project, and there are some really great accounts out there, and by ‘great’ I mean ‘scary’.
Here’s a key passage from Murphy and Mitchell (1974: p. 95):
V. salvator uses the tail to strike repeatedly in combination with biting for defense…Captive Varanus (varius, spenceri, mertensi, and salvadorii) use the tail for defense, but only salvadorii appears to aim directly for a handler’s eye. An adult male V. salvadorii accurately struck the senior author’s eye with the tip of the tail as he was attempting to maneuver the lizard. On many subsequent occasions, the monitor tried to strike the eye of the handler with accuracy.
Not being a monitor expert, I was initially thrown by the V. salvator/V. salvadorii issue. V. salvator is the water monitor, V. salvadorii is the crocodile monitor. Both get pretty darned big; Wikipedia lists 3.21 m (10.5 ft) for V. salvator and 2.44-3.23 m (8.0-10.6 ft) for V. salvadorii.
Anyway, I’d heard of lots of anecdotal reports of lizards from many clades using their tails to lash at rivals, predators, or handlers, but I’d never read about a lizard aiming directly for the target’s eyes. It immediately made me think about (1) sauropod tails, especially the whip-lash tails of flagellicaudan diplodocoids and at least some titanosaurs (Wilson et al. 1999), and (2) the supraorbital crests and ridges in many theropods, especially big Morrison forms like Allosaurus and Ceratosaurus. Of course, supraorbital crests in theropods could serve many functions, including shading the eyes and social and sexual display, but it’s interesting to speculate that they might have had a defensive function as well. Has anyone ever proposed that in print?

Diplodocus longus USNM 10865, from Gilmore (1932: plate 6)
Most of the papers that pooh-pooh the use of whiplash tails in defense (e.g., Myhrvold and Currie 1997) argue that the tail-tip would be too small to do any serious damage to a multi-ton attacker, and too fragile to survive an impact. This seems wrong-headed to me, like arguing that unless you find putative animal weapons broken and caked in their adversaries’ blood, they aren’t used as weapons. A structure doesn’t have to do lethal damage or any damage at all to serve as a weapon, as long as it dissuades a predator from attacking. I’d think that getting hit in the eye by a 35-foot bullwhip might convince an allosaur to go have a look at Camptosaurus instead.
Now, one could argue that if the whip-lash doesn’t do any serious damage, predators will learn to blow them off as dishonest signals (we’re assuming here that having your eye possibly knocked out doesn’t count as ‘serious damage’ to an allosaur). But it’s not like the whiplash was the only weapon a diplodocid could bring to bear: the proximal tail could probably deliver a respectable clobberin’, and then there’s the zero fun of being stomped on by an adversary massing a dozen tons or more. In that sense, the whip-lash is writing checks the rest of the body can certainly cash. It’s saying, “Getting hit with this will be no fun, and if that isn’t enough, there’s plenty more coming.”
All of this is leaving aside more obvious defensive adaptations of the tail in Shunosaurus, maybe Omeisaurus and Mamenchisaurus, and probably Spinophorosaurus (although I’d feel better about Spinophorosaurus if the association of the spikes and the tail was more secure). I suspect that all sauropod tails were useful in defense, but only some sauropod taxa used that behavior enough for a morphological enhancement (club, spikes, whiplash) to have evolved. Similarly, common snapping turtles, Chelydra serpentina, will wiggle their unspecialized tongues to attract fish (I’ve witnessed this myself in captive specimens) but lack the worm-shaped tongue lure found in the more ambush-specialized alligator snappers, Macrochelys temminckii. On reflection, there are probably few morphological changes in evolution that aren’t preceded by behavior. Not in a Lamarckian sense, just that certain variations aren’t useful unless the organism is already (suboptimally) performing the relevant function.
Bonus observation: Mike noted back when that Shunosaurus and Varanus retain complex caudal vertebrae all the way out to the end. Since in this case ‘complex’ means ‘having processes that muscles can attach to’, maybe that has something to do with keeping up relatively fine motor control in your bad-guy-whomping organ. Would be interesting to compare caudal morphology between tail-whomping lizards and committed caudal pacifists (assuming we can find any of the latter that we’re certain about – maybe tail-whomping just doesn’t get used very often in some taxa, like those that have caudal autotomy). Anyone know anything about that?
References
- Murphy, J. B., & Mitchell, L. A. (1974). Ritualized combat behavior of the pygmy mulga monitor lizard, Varanus gilleni (Sauria: Varanidae). Herpetologica, 90-97.
- Myhrvold, N. P., & Currie, P. J. (1997). Supersonic sauropods? Tail dynamics in the diplodocids. Paleobiology, 23(4), 393-409.
- Wilson, J. A., Martinez, R. N., & Alcober, O. (1999). Distal tail segment of a titanosaur (Dinosauria: Sauropoda) from the Upper Cretaceous of Mendoza, Argentina. Journal of Vertebrate Paleontology, 19(3), 591-594.
A permanent home for the SVP’s Aetogate documents
February 17, 2015
It’s hard to believe it’s been nearly seven years since the “resolution”, if you want to call it that, of Aetogate, the aetosaur plagiarism-and-claim-jumping scandal.
I was contacted privately today by someone wanting to know if I had copies of the SVP’s documents published in response to this. I didn’t — and the documents are hard to find since they have been moved at least twice from their original addresses on the SVP site. They can no longer be found by navigating that site — at least, not by me or my correspondent. (Searching doesn’t help either.)
But by suitable application of Google-fu I did manage to dig out the documents, in their third (at least) home. So I thought it would be useful to make permanent copies here, in case they vanish completely next time the SVP changes things around. Here they are:
- Statement from the Executive Committee about the allegations of unethical conduct from J. Martz, W. Parker, M. Taylor and M. Wedel against S. Lucas, A. Hunt, A. Heckert, and J. Spielmann.
[SVP copy] [local copy] - Best practices from the Ethics Education Committee regarding research, publication, and museum work.
[SVP copy] [local copy] - Addendum to Executive Committee’s Final Statement Concerning Allegations From Martz, Parker, Wedel and Taylor.
[SVP copy] [local copy]
(In case anyone’s forgotten, I was not impressed by these documents. At all.)
The Titanic was sunk by an Apatosaurus cervical
February 13, 2015
According to Rare Historical Photos from the 1860s to the 1960s, this is the iceberg that sank the Titanic:

photo of the iceberg that sunk the Titanic, taken the morning of April 15, 1912 from board of the ship “Prinz Adalbert”, before knowing the Titanic had sunk. The smear of red paint along the base of the berg (bottom right) prompted the chief steward to take the picture.
Clearly this was no iceberg, but a gigantic Apatosaurus vertebra, most of it hidden under water. Here is an artist’s impression:

They get everywhere, don’t they?
The planned vandalism of the Natural History Museum: a modest proposal
February 10, 2015
Go to Google and do a picture search for “natural history museum”. Here are the results I get. (I’m searching the UK, where that term refers to the British museum of that name — results in the USA may very.)

In the top 24 images, I see that half of them are of the building itself — rightly so, as it’s a beautiful and impressive piece of architecture that would be well worth visiting even if it was empty. Of the rest, ten are of specimens inside the museum: and every single one of them is of the Diplodocus in the main hall. (The other two photos are from the French natural history museum, so don’t really belong in this set. Not coincidentally, they are both primarily photos of the French cast of the same Diplodocus.)
The NHM’s Diplodocus — I can’t bring myself to call it “Dippy” is the icon of the museum. It’s what kids go to see. It’s what the museum used as the basis of the logo for the 2005 SVPCA meeting that was held there. It’s essentially the museum mascot — the thing that everyone thinks of when they think of the NHM.
And rightly so: it’s not just a beautiful specimen, it’s not just sensational for the kids. As the first cast ever made of the Carnegie specimen CM 84, it’s a historically important object in its own right. It was the first mounted Diplodocus ever, being presented in 1905 before the the original material was even on display in Pittsburgh.

As a matter of fact, this cast was the very first mounted sauropod to be publicly displayed: that honour is usually given to the AMNH Apatosaurus, but as museum-history expert Ilja Nieuwland points out:
The London ‘Dippy’ was in fact the first sauropod on public display, if only for three days in early July of 1904, in the Pittsburgh Exposition Society Hall.
There you have the Natural History Museum Diplodocus: the symbol of the museum, an icon of evolution, a historical monument, a specimen of great scientific value and unparalleled symbolism.
So naturally the museum management want to tear it down. They want to convert the Diplodocus hall into a blue whale hall. Because the museum doesn’t already have a blue whale hall.
Or, no — wait — it does already have a blue whale hall. That’s it. That’s what I meant to say. And very impressive it is, too.

I don’t mind admitting that the whale hall is my second favourite room in the museum. Whenever I go there as a tourist (rather than as a scientist, when I spend all my time in the basement), I make sure I see it. It’s great.
The thing is, it’s already there. A museum with a whale hall does not need another whale hall.
Obviously anticipating the inevitable outcry, the museum got all its ducks in a row on this. They released some admittedly beautiful concept artwork, and arranged to have opinion pieces written in support of the change — some by people who I would have expected to know better.
One of the more breathtaking parts of this planned substitution is the idea that Diplodocus is no longer relevant. The NHM’s director, Sir Michael Dixon says the change is “about asking real questions of contemporary relevance”. He says “going forward we want to tell more of these stories about the societally relevant research that we do”. This “relevance” rhetoric is everywhere. The museum “must move with the times to stay relevant”, writes Henry Nicholls in the Guardian.
There was a time when Diplodocus was relevant, you know: waaay back in the 1970s. But time has moved on, and now that’s 150,000,035 years old, it’s become outdated.
Conversely, the rationale for the whale seems to be that they want to use it as a warning about extinction. But could there ever be a more powerful icon of extinction than a dinosaur?
The thing is, the right solution is so obvious. Here’s what they want to do:

Clearly the solution is, yes, hang the whale from the ceiling — but don’t remove the Diplodocus. Because, seriously, what could be a better warning about extinction than the juxtaposition of a glorious animal that we lost with one that we could be about to lose?
All this argument about which is better, a Diplodocus or a blue whale: what a waste of energy. Why should we have to choose? Let’s have both.
I’ve even had an artist’s impression made, at great expense, to show how the combination exhibit would look. Check it out.

(If anyone would like to attempt an even better rendering, please by my guest. Let me know, and I’ll add artwork to this page.)
So that’s my solution. Keep the museum’s iconic, defining centrepiece — and add some more awesome instead of exchanging it. Everyone wins.
Can it be? Even more non-dinosaurian epipophyses? Yes, and this time they’re non-ornithodiran!
February 8, 2015
Having given pterosaurs all the glory in two earlier posts, it’s time to move yet further away from the sauropods we know and love, and look at epipophyses outside of Ornithodira.
Here, for example, is the basal archosauriform Vancleavea. (Thanks to Mickey Mortimer, whose a comment on an earlier post put us onto this, and various other candidate epipohysis-bearers which we’ll see below.)
Here is a pair of Vancleavea cervical vertebrae:

Nesbitt et al. (2009: fig. 11A). Vertebrae of Vancleavea campi. Two articulated cervical vertebrae (PEFO 33978) in left lateral view.
No ambiguity here: the epipophysis is even labelled.
But we can find epipophyses even outside Archosauriformes. Here, for example, is the the rhynchosaur Mesosuchus:

Dilkes (1998: fig. 7A). Mesosuchus browni. Holotype SAM 5882. Partial skull and jaws and cervical vertebrae in left lateral view.
Check out the rightmost vertebra (C7), clicking through for the full resolution if necessary. There is a definite eminence above the postzyg, separated from it by a distinct groove. Unless the drawing is wildly misleading, that is a definite epipophysis, right there.
But even more basal archosauromorphs have epipophyses. Check out Teraterpeton, described by Hans-Dieter Sues in 2003:

Sues (2003: figure 7). Teraterpeton hrynewichorum, NSM 999GF041 (holotype), cervical and anterior dorsal vertebrae and ribs, associated with right scapula (sc), ?clavicles (cl?), ?interclavicale (ic?), and incomplete right humerus (h), in right lateral view. Scale bar = 1 cm. a.p., accessory process above postzygapophysis; ax, axis; c3, c4, cervical vertebra 3 and 4, respectively; t, displaced tooth.
This is another one where the epipophysis is labelled (though not recognised as such — it’s just designated an “accessory process”).
Can we go yet more basal? Yes we can! Here are cervicals 2 and 3 of the trilophosaur Trilophosaurus (in an image that I rearranged and rescaled from the published original for clarity):

Spielmann et al. (2008: figure 30, rearranged). Cervical vertebrae 2-3 (i.e. axis and C3) of Trilophosaurus buettneri TMM 31025-140. Top row: right lateral. Second row: dorsal, with anterior to the left. Third row, left to right: anterior, left lateral, posterior. Bottom row: ventral, with anterior to the left.
The parts of this image to focus on (and you can click through for a much better resolution) are the postzyg at top right of the left-lateral view, which has a distinct groove separating the zygapophyseal facet below from the epipohysis above; and the posterior view, which also shows clear separation on both sides between these two structures.
While we’re playing with trilophosaurs here’s here’s another one (probably), Spinosuchus:

Spielmann et al. (2009: figure 3N). Spinosuchus caseanus holotype UMMP 7507, 5th cervical vertebra in left lateral view.
Again, the groove separating postzygapophyseal facet from epipophysis (at top right in the image) is clear.
But there’s more! Even the protorosaurs, pretty much the most basal of all archosauromorphs, have convincing epipophyses. Here are two that I found in Dave Peters’ post from two years ago, which I only discovered recently. [Here I must insert the obligatory disclaimer: while Dave Peters is a fine artist and has put together a really useful website, his ideas about pterosaur origins are, to put it mildly, extremely heterodox, and nothing that he says about phylogeny on that site should be taken as gospel. See Darren’s write-up on Tet Zoo for more details.]
Dave shows some probable, but not super-convincing epipophyses in the protorosaur Macrocnemus (shaded purple here) …

Cervicals 1-6 of the protorosaur Macrocnemus, modified from an uncredited image on Dave Peters’ site. Postzygapophyses in yellow, epipophyses in purple.
… and some much more convincing epipophyses in the better known and more spectacular protorosaur Tanystropheus:

Unspecified single cervical of Tanystropheus, from Dave Peters’ site. Postzygapophysis in yellow, epipohysis in purple.
Frustratingly, Dave doesn’t attribute these images, so I don’t know where they’re originally from (unless they’re his own artwork). Can anyone enlighten me? There’s a nice illustration in figure 57 of Nosotti’s (2007) epic Tanystropheus monograph that is at least highly suggestive of epipophyses:

Nosotti (2007:figure 57). Reconstruction of an anterior cervical vertebra (A) and of a mid-cervical vertebra (B) in small-sized specimens of Tanystropheus longobardicus. Left lateral view. Not to scale. Watercolor: Massimo Demma. Abbreviation pzp = postzygapophyseal process.
But it’s not as good as the one Peters used, as that one shows a distinct notch between postzyg and epipophysis, so I’d like to track that down if I can.
With this, I believe I am done on cataloguing and illustrating epipophyses, unless something dramatic turns up. (For example, this commenter thinks that nothosaurs have epipophyses, but I’ve not been able to verify that.) Here’s what we’ve found — noting that we’ve illustrated epipophyses on every taxon on this tree except Crocodylia:

So it seems that epipophyses may well be primitive at least for Archosauromorpha — which implies that they were secondarily lost somewhere on the line to modern crocs.
With this lengthy multi-part digression complete, hopefully, we’ll get back to sauropods next time!
References
- Dilkes, David W. 1998. The Early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles. Philosophical Transactions of the Royal Society of London B 353:501-541.
- Kellner, Alexander W. A., and Yukimitsu Tomida. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum monographs, Tokyo, 17. 135 pages.
- Nesbitt, Sterling J., Michelle R. Stocker, Bryan J. Small and Alex Downs. 2009. The osteology and relationships of Vancleavea campi (Reptilia: Archosauriformes). Zoological Journal of the Linnean Society 157:814-864.
- Nosotti, Stefania. 2007. Tanystropheus longobardicus (Reptilia, Protorosauria): re-interpretations of the anatomy based on new specimens from the Middle Triassic of Besano (Lombardy, Northern Italy). Memorie della Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 35(III). 88pp.
- Spielmann, Justin A., Spencer G. Lucas, Larry F. Rinehart and Andrew B. Heckert. 2008. The Late Triassic Archosauromorph Trilophosaurus. New Mexico Museum of Natural History and Science Bulletin 43.
- Justin A. Spielmann, Spencer G. Lucas, Andrew B. Heckert, Larry F. Rinehart and H. Robin Richards III. 2009. Redescription of Spinosuchus caseanus (Archosauromorpha: Trilophosauridae) from the Upper Triassic of North America. Palaeodiversity 2:283-313.
- Sues, Hans-Dieter. 2003. An unusual new archosauromorph reptile from the Upper Triassic Wolfville Formation of Nova Scotia. Canadian Journal of Earth Science 40:635-649.
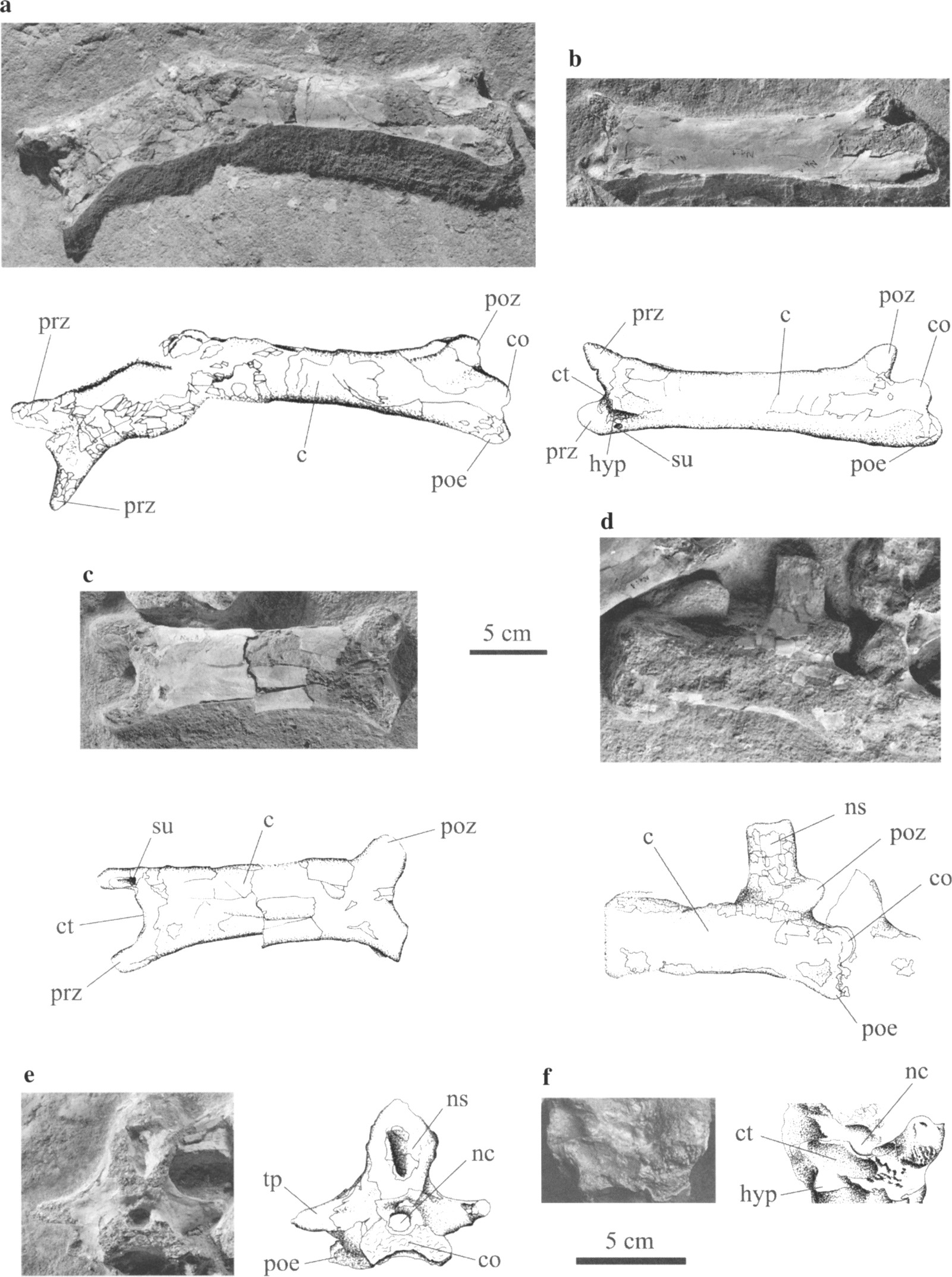
What’s that you say? No epipophyses in titanosaurs? Oh really?
February 6, 2015
This just in, from Zurriaguz and Powell’s (2015) hot-off-the-press paper describing the morphology and pneumatic features of the presacral column of the derived titanosaur Saltasaurus. (Thanks to Darren for bringing this paper to my attention.)
Now, as everyone knows, titanosaurs don’t have epipophyses. In fact, they’re the one major sauropod group where Matt has not observed them.
Until today.

Zurriaguz and Powell (2015:figure 3B). Anterior cervical vertebra PVL 4017-3 of Saltasaurus loricatus, in dorsal view (rotated 90° from the paper)
Look at the left postzygapophysis, at top left of this image. Doesn’t that look like there’s a distinct rounded eminence sticking out towards the camera?
No? Not convinced? All right, then, how about this?

Zurriaguz and Powell (2015:figure 4B). Mid-anterior cervical PVL 4017-138 of Saltasaurus loricatus in right lateral view.
This time, look at the right postzyg (again at top left in the image). Doesn’t that look like there are two separate bony structures up there separated by a notch? A postzygapophyseal facet below, and an epipophysis above? Right?
Huh? What’s that? Just damage, you say?
All right. Let’s bring out the smoking gun.

Zurriaguz and Powell (2015:figure 5). Last anterior cervical vertebra (PVL 4017-5) of Saltasaurus loricatus in right lateral view. (Ignore the inset square for our purposes: it’s in the original.)
Again up at top left, we seem to have a clear case of a ventrally directed postzygapophyseal facet surmounted by a separate eminence which can only be an epipophysis. It even seems to be roughened for tendon attachment.
What does this mean? Only the same thing we said last time: The more we look for epipophyses, the more we find them. Amazing how often that turns out to be true of various things.
We seem to be headed towards the conclusion that epipophyses, while never ubiquitous, pop up in all sorts of places scattered all across the ornithodiran tree, encompassing birds, other theropods, sauropods, prosauropods, several groups of ornithischians, and both pterodactyloid and “rhamphorhynchoid” pterosaurs.
But what about outside Ornithodira?
Can we find epipophyses even out there, in the wilderness?
Stay tuned!
References
The equivocal epipophyses of Cf. Quetzalcoatlus
February 5, 2015
It’s well known that there is good fossil material of the giant azhdarchid pterosaur Quetzalcoatlus out there, but that for various complicated reasons it’s yet to be published. But as part of our ongoing quest for pterosaur epipophyses, I have obtained these photos of a pretty well preserved single cervical, probably C3, which is either Quetzalcoatlus or something pretty darned close.

TMP 1992.83.7, Cf. Quetzalcoatlus, cervical 3. Top, dorsal view; bottom, ventral view. Anterior is to the left. Scale bar = 10 cm. Click through for high resolution.
My thanks go, in chronological order, to Rob Knell of QMC for taking the photos; to Don Brinkman for permission to share them publicly; and to Mike Habib (the USC one, not the Elsevier one) for passing them on to me. (The composition is my own work, which anyone is free to reuse so far as I’m concerned.)
Here’s what Mike Habib says about the specimen:
… well preserved TMP azhdarchid cervical vertebra. It is likely a CIII vert, and appears to be from an animal very similar to the small morph of Quetzalcoatlus in overall morphology. The associated humerus is just about an exact match. This cervical, however, does not quite match the proportions of any of the Q. sp. cervical verts, though that’s not a surprise given that the animals come from different horizons. There is a much larger, but poorly preserved cervical vert at the TMP as well (a Q. northropi sized animal, give or take).
Here are Mike’s measurements:
- maximum length: 142.2 mm
- minimum mediolateral breadth: 39.3 mm
- minimum dorsoventral breadth: 27.4 mm
- midshaft mediolateral breadth 40.0 mm
- midshaft dorsoventral breadth 27.0 mm
- mediolateral breadth across prezygapophyses: 65.6 mm
- mediolateral breadth across postzygapophyses: 68.5 mm
- dorsoventral breadth at postzygapophyses: 35.8 mm
(I mean those are the measurements that Mike provided for the vertebra, not the measurements of Mike himself. He’s much bigger than that.)
So does this specimen have epipophyses? Frustratingly, there don’t seem to be lateral or posterior-view photos, so it’s very hard to tell from these dorsal and ventral ones. Happily, the same specimen was illustrated and briefly described by Godfrey and Currie (2005:294-299), along with several other less well-preserved cervicals — so we do have drawings of these other views:

Godfrey and Currie (2005:figure 16.1). Azhdarchid cervical vertebra (TMP 92.83.7) in (A) dorsal, (B) left lateral, (C) ventral, (D) anterior, (E) posterior, and (F) posterodorsal views. Abbreviations: hyp, hypapophysis; nc, neural canal; pn, pneumatopore; prz, prezygapophysis.
(The specimen number given here is slightly different from that given for the photos, but matches the label in the ventral-view photo. I assume that the leading “93” part of the specimen number is a year, and that it’s sometimes but not always given in four digits.)
The text of the description does not mention epipophyses, and skips very lightly over the whole postzygapophyseal area. But figures 16.1B (lateral) and 16.1E (posterior) both seem to show distinct bulbous eminences well above the postzygapophyseal facets. I think these have to be epipophyses. So Mark Witton’s caution not to write off azhdarchid epipophyses on the strength of their apparent absence in Phosphatodraco proves well-founded.
What is the moral here?
The more we look for epipophyses, the more we find them.
Which will be strangely familiar to anyone who remembers our experience with caudal pneumaticity in sauropods, which was: the more we looked for it, the more we found it.
If we have an SV-POW! motto (other than “sauropods are awesome”, of course), it’s “Measure your damned dinosaur!“. But if we had a third motto, it would be like unto it: look at your damned dinosaur. Or pterosaur, as the case may be. The odds are, you’ll see things you weren’t expecting.
Many thanks for the various people who chipped in, both in comments on the last post and in this thread on twitter, where I asked a bunch of pterosaur experts for their thoughts on epipophyses in pterosaurs. I now know more than I previously knew about epipophyses outside of Sauropoda — and especially outside Dinosauria. I’ll try to credit everyone who contributed.
Occasional SV-POW!sketeer Darren Naish claims that according to the literature, ornithischians lack epipophyses — something that we’ve seen is untrue. I never got references out of him, though. Can anyone point me to the guilty literature?
Darren also gave me the rather cryptic instruction “Look at Anhanguera monographs. Sorry, can’t check myself.” Like something from a spy novel. Checking out Kellner and Tomida (2000), I found their illustration of the Anhanguera atlas/axis complex, in figure 14B, suggestive:

It took me a while to figure this out, but I think this is showing the first three cervicals, not two: the atlas is tiny, and is smushed onto the front of the axis; C3 is shown, but only in outline, and is ignored in the caption.
As labelled, the postzygapophyseal facet of the axis is tiny — and there’s a definite protuberance above it, which can only be an epipophysis. But we’d need photos to be confident. The good news is that there is a photo in the paper — part A of the same figure. But the bad news is that here’s how it looks in my scan:

Not so helpful. If anyone has a good scan — or better still an original photo — I’d like to see it.
Darren also commented “Most big pterosaurs lack epipophyses. Ornithocheirids may be the exception”, but there his hints dried up. Mark Witton cautioned me: “Not sure for azhdarchids. Well preserved verts have reduced features, but not entirely absent as badly preserved verts suggest.” So perhaps the Phosphatodrado vertebrae in the last post are not so compelling as they seem.
Liz Martin suggested “off the top of my head you could check Wellnhofer papers. 1991 and 1985 I think show verts.” But I couldn’t find any vertebrae in the only Wellnhofer (1985) that I have; and there are at least three Wellnhofer publications from 1991, which I’ve not checked yet. Any more guidance, anyone?
So how widespread are epipohyses? Brusatte et al. (2010:73) gave “Epipophyses on the cervical vertebrae” as a synapomorphy diagnosing Dinosauria:
2.4.1.4. Epipophyses on the cervical vertebrae. Epipophyses are projections of bone, likely for muscle and ligament attachment, which protrude from the dorsal surfaces of the postzygapophyses of the cervical vertebrae. All basal dinosaurs possess epipophyses (Langer and Benton, 2006), although the size, shape, length, and projection angle of these processes vary considerably (e.g., compare Coelophysis (Colbert, 1989) with the more derived theropod Majungasaurus (O’Connor, 2007)). Basal ornithischians (e.g., Heterodontosaurus) only have epipophyses on the anterior cervical vertebrae, whereas saurischians have epipophyses in nearly all cervical vertebrae (Langer and Benton, 2006). Epipophyses are not present in the closest relatives of dinosaurs (e.g., Marasuchus, Silesaurus), but are present in some crurotarsans (e.g., Lotosaurus and Revueltosaurus).
It’s surprising that they’d mention dinosaurs and croc-line archosaurs, but overlook pterosaurs, which are phylogenetically bracketed by that group. But there’s lots of useful detail to follow up in the citations, which I’ll be doing soon.
So: moving down the tree from Sauropoda, we see epipophyses:
- often but not always in sauropods
- rarely in basal sauropodomorphs
- often, maybe always, in theropods
- intermittently but not infrequently in ornithischians
- in at least some basal dinosauriforms
- in some groups of pterosaurs but not others
- in at least some croc-line archosaurs — but not, for example, in alligators.
Does anyone know of epipophyses outside Archosauria?
We seem now to be stumbling towards a conclusion of sorts, which is that epipophyses seem to be rather phylogenetically labile, coming and going within numerous lineages. As with so many vertebral features, they also vary with serial position, which complicates matters; and, I dare guess, with ontogeny.
I’ve not been able to locate any publications that are specifically about epipophyses (just lots that mention them in passing). Does anyone know of such a thing?
References
- Brusatte, Stephen L., Sterling J. Nesbitt, Randall B. Irmis, Richard J. Butler, Michael J. Benton, and Mark A. Norell. 2010. The origin and early radiation of dinosaurs. Earth-Science Reviews 101(1):68-100.
- Kellner, Alexander W. A., and Yukimitsu Tomida. 2000. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), Northeastern Brazil. National Science Museum monographs, Tokyo, no. 17. 135 pages.
- Wellnhofer, Peter. 1985. Neue pterosaurier aus der Santana-Formation (Apt) der Chapada Do Araripe, Brasilien. Paläontographica A 187:105-182. [in German]

{kind=link}
{kind=link}
{kind=link}